

- Гистология
- Гладкая (висцеральная) мускулатура
- Историческая справка
- Молекулярные механизмы сокращения скелетной мышцы
- Ответ мышц на физическую нагрузку
- Отдых
- Работа скелетной мышцы
- Саркомер при сокращении мышцы
- Сердечная поперечнополосатая мышечная ткань
- Скелетная (поперечнополосатая) мышечная ткань
- Сокращение
- Состав саркомера
- Сравнительная структура
- Структурная организация скелетной мышцы
- Фазы и режимы сокращения скелетной мышцы
- Физиологические свойства мышц
Гистология

Гистологические срезы мышцы показывают анатомические особенности саркомеров. Толстые нити, состоящие из миозина, видны и представлены как полоса А саркомера.
Тонкие нити, состоящие из актина, связываются с белком в Z-диске (или Z-линии), называемым альфа-актинином, и присутствуют на всей длине I-полосы и части A-полосы.
Область, в которой перекрываются толстые и тонкие волокна, выглядит плотно, так как между ними мало места. Эта область, где тонкие и толстые волокна перекрываются, очень важна для сокращения мышц, так как именно здесь начинается движение волокна.
Тонкие волокна не полностью переходят в полосы А, оставляя центральную область полосы А, которая содержит только толстые волокна. Эта центральная область полосы A кажется немного светлее, чем остальная часть полосы A, и называется зоной H.
В центре зоны H есть вертикальная линия, называемая линией M, где дополнительные белки удерживают вместе толстые волокна.
Основные компоненты гистологии саркомера резюмируются ниже:
Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках
желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все
остальные клетки.
Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы
внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют
такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой.
К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.
Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Историческая справка
В 1954 году ученые опубликовали две новаторские статьи, описывающие молекулярную основу сокращения мышц. В этих работах описывалось положение миозиновых и актиновых филаментов на разных стадиях сокращения мышечных волокон и предлагалось, как это взаимодействие вызывает сократительную силу.
Используя микроскопию высокого разрешения, А. Ф. Хаксли и Р. Нидерберг (1954) и Х. Э. Хаксли и Дж. Хансон (1954) наблюдали изменения в саркомерах по мере сокращения мышечной ткани. Они заметили, что одна зона повторного саркомера, «полоса А», оставалась относительно постоянной по длине во время сокращения.
Эти наблюдения побудили их предложить теорию скользящих нитей, в которой говорится, что скольжение актина относительно миозина вызывает напряжение мышц. Поскольку актин привязан к структурам, расположенным на боковых концах каждого саркомера, называемых z-дисками или «z-полосами», любое сокращение длины нити актина приведет к сокращению саркомера и, следовательно, мышцы.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
• Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
• Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
• Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са (рис. 2, В).
• Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
«Вверх»
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной
массы нарастает.
В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной
атрофии (греч. а – «не» trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.
Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в
размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление.
Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае
гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).
Отдых
В покое головка миозина связана с АТФ молекула имеет низкоэнергетическую конфигурацию и не может получить доступ к сайтам связывания поперечного мостика на актине. Однако головка миозина может гидролизовать АТФ в аденозиндифосфат (АДФ и неорганический фосфат-ион.
Часть энергии, высвобождаемой в этой реакции, изменяет форму миозиновой головки и приводит ее к высокоэнергетической конфигурации. В процессе связывания с актином головка миозина высвобождает АДФ и неорганический фосфат-ион, меняющий свою конфигурацию обратно на низкоэнергетическую.
Миозин остается прикрепленным к актину в состоянии, известном как строгость, пока новый АТФ не свяжет миозиновую головку. Это связывание АТФ с миозином высвобождает актин за счет диссоциации поперечного мостика. Миозин, связанный с АТФ, готов к следующему циклу, начиная с гидролиза АТФ.
Полоса А видна в виде темных поперечных линий на миофибриллах; I-полоса видна как слегка окрашивающиеся поперечные линии, а Z-линия видна как темные линии, разделяющие саркомеры на уровне светового микроскопа.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
• числом ДЕ, участвующих в сокращении;
• частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
• динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
• статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
• динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
• изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
• изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
• ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма.
«Вверх»
Саркомер при сокращении мышцы
• Миозин II представляет собой мотор, обеспечивающий сокращение мышц
• Актин и миозин II являются основными компонентами саркомера, основной единицей сокращения поперечнополосатых мышц
Мышцы представляют собой ткань, способную к сокращению, которая обеспечивает движение тела и подвижность органов. В соответствии с внешним видом сократительных волокон, мышцы можно подразделить на две основных группы: поперечнополосатые и гладкие. Волокна поперечнополосатых мышц называются так потому, что при большом увеличении можно видеть их характерную исчерченность. К этой группе относятся скелетные и сердечная мышцы.
Скелетные мышцы обеспечивают подвижность скелета, а сердечная сокращение сердца. Волокна гладких мышц не исчерченные и имеют веретенообразную форму. Эти мышцы образуют стенки органов, например мочевого пузыря, кровеносных сосудов и желудочно-кишечного тракта.
Белковый мотор, который обеспечивает сокращение мышц, относится к семейству миозина II. Этот миозин принадлежит к числу наиболее распространенных белков позвоночных, он легко выделяется и поэтому является одним из хорошо изученных белков.
Миозин II представляет собой гексамерный комплекс,
состоящий из двух тяжелых цепей и двух пар различных легких цепей.
Эти комплексы собираются в биполярные толстые филаменты.
Дистальный участок хвоста мышечного миозина II связан с другими молекулами миозина II, образуя волокна, состоящие примерно из 300 молекул миозина. Эти волокна биполярны, и моторные домены всех миозинов направлены от центральной зоны. Эти волокна называются толстые биполярные филаменты.
Поперечнополосатые мышцы состоят из пучков мышечных волокон. Последние представляют собой крупные многоядерные клетки, длина которых варьирует от нескольких миллиметров до нескольких сантиметров, а диаметр составляет 20-100 мкм. Рисунок ниже показывает, что каждое мышечное волокно состоит более чем из 1000 миофибрилл, которые представляют собой палочковидные органеллы, способные к сокращению.
Саркомеры представляют собой основные сокращающиеся единицы поперечнополосатых мышц; при сокращении и расслаблении мышцы их длина изменяется. Как показано на рисунке ниже, саркомеры содержат толстые филаменты, состоящие, главным образом, из биполярных филаментов миозина II, и тонких филаментов, которые содержат актиновые нити и регуляторные белки.
Актиновые филаменты прикрепляются к Z-диску и копируются за счет связывания с копирующим белком (CapZ), что предотвращает деполимеризацию актиновых филаментов. Заостренные концы актиновых филаментов ориентированы к центру саркомера и копированы тропомодулином.
Толстые филаменты находятся в центре, на М-линии, между Z-дисками. М-линия представляет собой структуру, состоящую из гибко связанных биполярных толстых филаментов; эти связи скрепляют толстые филаменты, гексагонально расположенные внутри.
Наряду с М-линией, белок филаментов, который называется титаном, образует эластичные связи между Z-дисками и миозиновыми волокнами. Титин обеспечивает центровку толстых филаментов в саркомере и действует как пружина, которая не допускает растягиваться саркомеру.
Тонкие и толстые филаменты переплетаются, образуя трехмерную решетчатую структуру. Поскольку саркомер биполярен, в обеих половинах миозиновые моторы по отношению к актину ориентированы одинаково. При сокращении моторные домены миозина толстых филаментов взаимодействуют с актином тонких филаментов.
При сокращении саркомер укорачивается за счет скольжения тонких и толстых филаментов относительно друг друга, что сближает соседние Z-диски к центру саркомера. По мере продвижения головок миозина к зазубренным концам актиновых филаментов, длина толстых и тонких филаментов остается постянной.
Ширина саркомера в расслабленной мышце позвоночных составляет около 3 мкм, а при сокращении она становится около 2,4 мкм.
В мышечном волокне укорачиваются группы, состоящие из тысяч саркомеров, что приводит к укорачиванию всей мышцы. Общая длина укороченного мышечного волокна определяется двумя факторами: длиной, на которую укорачивается каждый саркомер, и количеством саркомеров в группе. Процентная величина, на которую происходит укорачивание мышечных волокон, одна и та же, независимо от их длины.
Величина усилия, генерируемого саркомером, пропорционально числу акто-миозиновых взаимодействий в половине саркомера, а величина усилия, которое развивает мышчное волокно, пропорционально числу параллельно расположенных саркомеров. Таким образом, штангисты увеличивают свою силу за счет увеличения площади поперечного сечения мышц, а не за счет их длины.
Как показано на рисунке ниже, сокращение поперечнополосатых мышц регулируется с участием тропонин-тропомиозинового комплекса, который связан с актином в тонких филаментах. Молекулы тропомиозина представляют собой суперспирализованные полипептиды длиной 40 нм.
Она располагаются друг за другом вдоль актиновых спиралей. Тропонин представляете собой комплекс из трех различных белков: тропонина С, тропонина I и тропонина Т. Один комплекс связывается с тропомиозином так, что они располагаются вдоль тонких филаментов через 40 нм интервалы.
При низких концентрациях ионов кальция тропомиозин находится в таком состоянии, что пространственно блокирует сайт связывания миозина на актине, так что мышца расслабляется и гидролиз АТФ под действием миозина происходит очень медленно. За счет взаимодействия актина с миозином релаксированные саркомеры способны к пассивному растяжению, оказывая небольшое сопротивление.
Нервные импульсы вызывают выход кальция в цитозоль из саркоплазматического ретикулума. Последний представляет собой органеллу, которая депонирует кальций в мышце. Повышение уровня кальция в цитозоле приводит к его связыванию с тропонином-С и к конформационным изменениям в молекуле белка.
Скелетные мышцы состоят из мышечных волокон, содержащих миофибриллы и представляющих собой удлиненные клетки.
Миофибриллы способны к сокращению и содержат повторяющиеся структурные единицы, называемые саркомерами.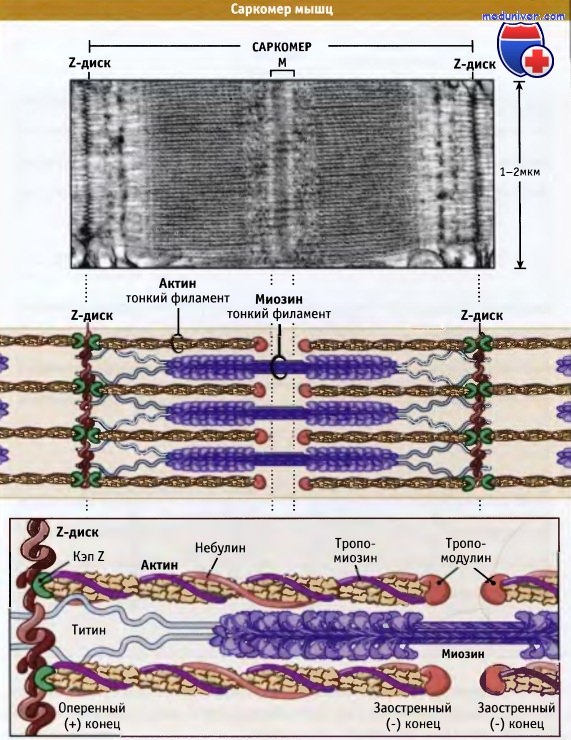 На конце каждого саркомера находится Z-диск, к которому через CapZ (кэпирующий белок) присоединяются актиновые филаменты.
На конце каждого саркомера находится Z-диск, к которому через CapZ (кэпирующий белок) присоединяются актиновые филаменты.
Толстые миозиновые филаменты соединяются с Z-диском через белок титин и вплетаются между актиновыми филментами.
Небулин проходит от Z-диска к тропомодулину. Однако как он связывает актин, пока неизвестно.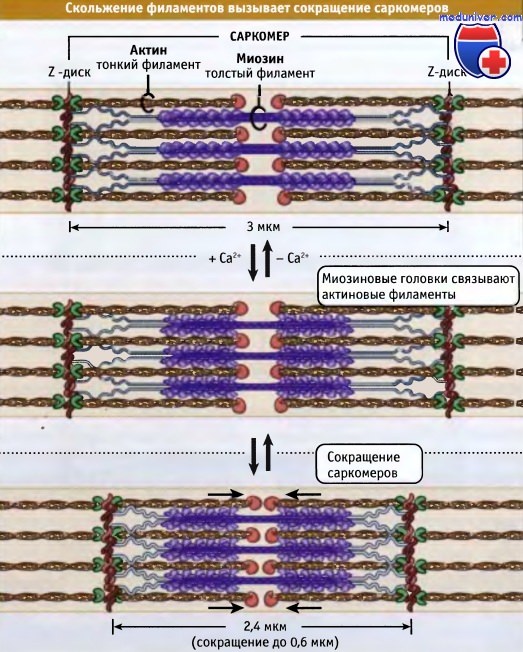 Сокращение мышцы происходит, когда толстые миозиновые филаменты присоединяются к актиновым и перемещают их таким образом,
Сокращение мышцы происходит, когда толстые миозиновые филаменты присоединяются к актиновым и перемещают их таким образом,
что Z-диски сдвигаются относительно друг друга. При этом длина саркомера уменьшается.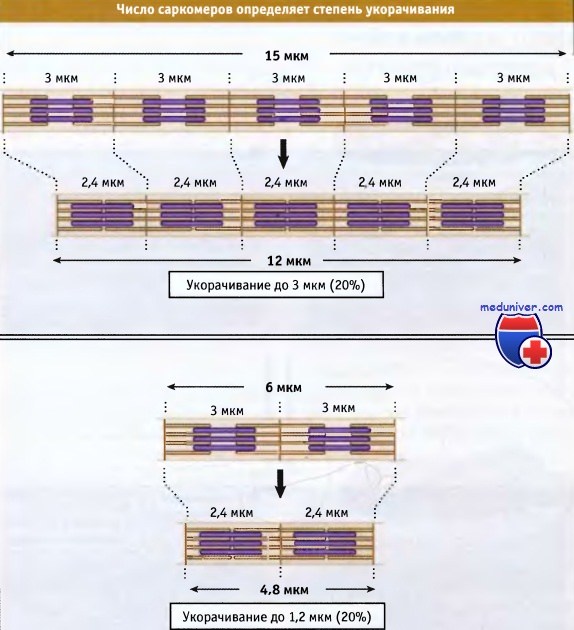 Чем длиннее миофибрилла (т. е. чем больше саркомеров), тем более она укорачивается при сокращении.
Чем длиннее миофибрилла (т. е. чем больше саркомеров), тем более она укорачивается при сокращении.
Однако в процентном отношении величина укорачивания не зависит от числа саркомеров.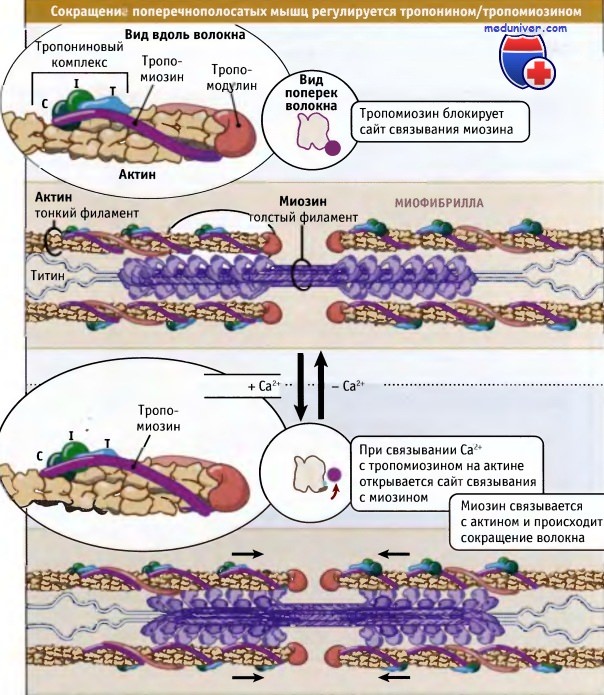 Сокращение поперечнополосатых мышц регулируется уровнем Са2 в цитозоле,
Сокращение поперечнополосатых мышц регулируется уровнем Са2 в цитозоле,
который определяет положение комплекса тропонин/тропомиозин относительно миозина и актина.
Источник
Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения
сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).
В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они
спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям
ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными
волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе plast — образованный) представляет совокупность слившихся клеток, имеет длину
от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).
Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос
на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего
все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Сокращение
Протеин тропомиозин покрывает миозин-связывающие участки молекул актина в мышечной клетке. Чтобы мышечная клетка сократилась, тропомиозин должен быть перемещен, чтобы раскрыть участки связывания на актине. Ионы кальция связываются с тропонин С молекулы (которые рассредоточены по всему белку тропомиозина) и изменяют структуру тропомиозина, заставляя его обнаруживать сайт связывания поперечного мостика на актине.
Концентрация кальция в мышечных клетках контролируется саркоплазматический ретикулум, уникальная форма эндоплазматический ретикулум в саркоплазма.
Мышечные клетки стимулируются, когда двигательный нейрон освобождает нейромедиатор ацетилхолин, который путешествует по нервномышечное соединение (синапс между конечным бутоном нейрона и мышечной клеткой). Ацетилхолин связывается с постсинаптическим никотиновый рецептор ацетилхолина.
Изменение конформации рецептора позволяет притоку натрий ионов и инициирование постсинаптического потенциал действия. Потенциал действия затем перемещается Т-канальцы (поперечные канальцы), пока не достигнет саркоплазматического ретикулума.
Здесь деполяризованная мембрана активирует потенциалзависимый Кальциевые каналы L-типа, присутствуют в плазматической мембране. Кальциевые каналы L-типа находятся в тесной связи с рианодиновые рецепторы присутствует на саркоплазматической сети.
Входящий поток кальция из кальциевых каналов L-типа активирует рианодиновые рецепторы для высвобождения ионов кальция из саркоплазматического ретикулума. Этот механизм называется кальций-индуцированное высвобождение кальция (CICR). Неизвестно, вызывает ли открытие рианодиновых рецепторов физическое открытие кальциевых каналов L-типа или присутствие кальция.
Сокращение мышц заканчивается, когда ионы кальция закачиваются обратно в саркоплазматический ретикулум, позволяя сократительному аппарату и, таким образом, мышечным клеткам расслабиться.
При сокращении мышцы полосы А не меняют своей длины (1,85 мкм в скелетных мышцах млекопитающих),[3] тогда как I-полосы и H-зона укорачиваются. Это приводит к сближению линий Z.
Состав саркомера
Чаще всего при описании состава саркомера указывается, что саркомер состоит из толстых и тонких филаментов, и приводится следующий рисунок (рис.1).
Однако этот рисунок не дает полное представление как о составе, так и о структуре саркомера. Не будем забывать, что саркомер – это объемный объект, маленький цилиндр.
Рис.1. Схема строения саркомера (G.H. Pollak, 1990) Длина саркомера ограничена Z-дисками. В середине саркомера располагается М-диск, который представляет собой «сцепление хвостов миозиновых молекул».
Если мысленно разрезать саркомер поперек, то в месте, где пересекаются толстые и тонкие филаменты, мы увидим следующую картину (рис.2).
Рис. 2. Решетчатая структура филаментов актина и миозина (H.E. Huxley, 1972)
Отчетливо видно, что каждый толстый филамент окружен шестью тонкими филаментами, то есть видно большое количество структурных элементов. Еще в 1972 году известный гистолог А.Н. Студитский выделил эти структурные элементы в составе саркомера. Для их описания он использовал следующее образное сравнение:
«На поперечных срезах в миофибриллах открывается картина взаимного расположения филамент, гексогональная упаковка, напоминающая связку карандашей, повернутую концами к зрителю». Из этого образного сравнения следует, что А.Н. Студитский сравнивает эти структурные элементы с карандашами, а саркомер, в свою очередь, представляет связку таких «карандашей».
В нашей статье (А.В. Самсонова, Г.А. Самсонов, 2022) мы предложили называть такие элементы, из которых состоят саркомеры – сотами по аналогии с пчелиными сотами.
Сот– структурная единица саркомера, в состав которой входит один толстый и 12 тонких филаментов, шесть из которых прикреплены к Z-диску с одной стороны от толстого филамента, а другие шесть – с другой стороны от толстого филамента (рис. 3).
Длина сота равна длине саркомера, так как длина сота ограничена с двух сторон Z-дисками. На рис. 3 представлены основные элементы, входящие в состав сота: один толстый филамент и шесть тонких филаментов, расположенных справа и слева от толстого филамента, а также часть Z – диска, к которому прикреплены тонкие филаменты.
Следует отметить, что состав и строение этого элемента значительно сложнее. Например, на рисунке не указано, что белок титин (тайтин) соединяет каждый толстый филамент с Z-дисками. Данная схема предназначена для того, чтобы выделить этот структурный элемент в составе саркомера.
Рис. 3. Схема строения сота. Толстый филамент, подобно грифелю карандаша расположен в середине, на ребрах карандаша расположены тонкие филаменты. Шесть тонких филаментов прикреплены к Z-диску с одной стороны от толстого филамента, а другие шесть – с другой стороны.
Установлено (Г. А. Самсонов, А.В. Самсонова, 2022), что в одном саркомере, имеющем диаметр 1 мкм находится более 1000 таких структурных элементов – сот. То есть в связке «карандашей» более 1000. Площадь одного сота равна 6,225 10 -4 мкм 2 .
В 2000 году математик Томас Хелс доказал, что шестиугольники, лежащие в основе пчелиных сот, лучше других геометрических фигур подходят для максимального использования пространства, при этом используется минимум строительного материала (воска). По-видимому, это свойство гексагональной структуры сотов также используется и при «строительстве» саркомеров.
Сравнительная структура
Структура саркомера влияет на его функцию несколькими способами. Перекрытие актина и миозина приводит к кривая длина-растяжение, который показывает, насколько саркомер сила выход уменьшается, если мышца растягивается так, что меньше поперечных мостиков может образовываться или сжиматься до тех пор, пока актиновые филаменты не будут мешать друг другу.
Длина актиновых и миозиновых нитей (взятая вместе как длина саркомера) влияет на силу и скорость — более длинные саркомеры имеют больше поперечных мостиков и, следовательно, большую силу, но имеют меньший диапазон укорочения. Позвоночные животные демонстрируют очень ограниченный диапазон длин саркомеров с примерно одинаковой оптимальной длиной (длина при пиковом напряжении длины) во всех мышцах человека, а также между видами.
Членистоногиетем не менее, демонстрируют огромные различия (более семи раз) в длине саркомеров как между видами, так и между мышцами у одного человека. Причины отсутствия значительной вариабельности саркомеров у позвоночных до конца не известны.[нужна цитата]
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые построены из повторяющихся в продольном направлении блоков (саркомеров).
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ). ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
«Вверх»
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
• латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
• фаза укорочения (около 50 мс);
• фаза расслабления (около 50 мс).
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
• Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
• При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
• При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды.
«Вверх»
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1.Сравнительная характеристика скелетных и гладких мышц
Пластичность гладких мышц проявляется в том, что они могут сохранять постоянный тонус как в укороченном, так и в растянутом состоянии.
Проводимость гладкой мышечной ткани проявляется в том, что возбуждение распространяется от одного миоцита к другому через специализированные электропроводящие контакты (нексусы).
Свойство автоматии гладкой мускулатуры проявляется в том, что она может сокращаться без участия нервной системы, за счет того, что некоторые миоциты способны самопроизвольно генерировать ритмически повторяющиеся потенциалы действия.
«Вверх»

